FECUNDACION
El desarrollo
de las técnicas
de fecundación in vitro –FIV – así como
las técnicas de
cultivo de embriones
preimplantados en mamíferos, han permitido comprender los
mecanismos celulares y moleculares
de la interacción gamética, que en último
llevan a la
fecundación
Para que tenga lugar la fecundación es necesario
que los gametos tanto femeninos como masculinos sufran un proceso de maduración
intracelular que los hace aptos para el intercambio de estructuras que darán
lugar a un nuevo individuo.

TRANSPORTE OOCITO

TRANSPORTE GAMETICO

Después de la ruptura folicular, el óvulo es recogido por la trompa, la cual entra en contacto con el ovario por medio de las contracciones miométricas, el músculo liso del mesosalpinx y los ligamentos tuboováricos.
Poco después alcanza la
ampolla tubárica sitio de la fecundación.
La vida fértil del óvulo eclosionado permanece
viable entre 12 a 24 horas después. Pierde su capacidad fecundante al llegar al
istmo del oviducto y es completamente infecundo al llegar al útero.
Todos los óvulos no fecundados se desintegran y
fagocitan dentro del útero.
El tiempo de transporte
del óvulo en el oviducto varía según
la especie y depende de :
· - Contracción
de la musculatura oviduto.
· - Corrientes
y contracorrientes de los líquidos en el
oviducto
· - Actividad
secretora de las células ciliadas influida por los estrógenos y progesterona
· - Hidrodinámica
y propiedades reológicas de los líquidos
oviductales.
Es
importante la sincronía entre la ovulación
y la inseminación con el fin de
lograruna pronta fecundación.
En monta
natural el envejecimiento del óvulo antes
de la penetración del
espermatozoide es bajo.
TRANSPORTE
DEL ESPERMATOZOIDE
Durante la eyaculación los
espermatozoides junto con el plasma seminal pasan por la uretra y a través de
movimientos peristálticos se liberan en el tracto genital femenino. La
eyaculación es el reflejo de expulsión de los espermatozoides y el plasma seminal
fuera del tracto reproductivo.
El reflejo eyaculatorio es el
resultado de la estimulación sensorial especialmente en el glande, lo que causa
contracciones musculares coordinadas. Una vez se introduce el pene en la vagina
se inicia el reflejo por impulsos que se transmiten del glande a través del
nervio púdico hasta la región lumbosacra de la médula espinal. Así el semen es
forzado a pasar a la uretra lo que induce la contracción de los músculos
uretrales, isquiocavernosos y bulboespongiosos. El eyaculado contiene, además,
las secreciones de las glándulas anexas (vesículas seminales, próstata,
glándulas bulbouretrales).
Luego de depositado el semen, los
espermatozoides están expuestos a una serie de circunstancias ambientales que
alteran significativamente su número y función.
Las pérdidas de espermatozoides en
el tracto genital femenino depende de la naturaleza física
del eyaculado y el sitio de la deposición. Los
espermatozoides se pierden en el tracto femenino
por transporte retrógrado y muchos son fagocitados por
los leucocitos, que desde el punto de vista
inmunológico actúan como cuerpos extraños, no
diferenciando entre vivos o muertos.

De hecho un leucocito puede fagocitar varios espermatozoides móviles. Además son importantes para prevenir infecciones del tracto genital.
El transporte de espermatozoides después de la
cópula puede dividirse en dos fases, una fase de
transporte rápido y una fase de transporte lento o
prolongado durante la fase rápida los mecanismos del tracto
genital permiten la llegada de los espermios al
oviducto sin la participación activa de estos.

Lo más importante del transporte es la fase de transporte lento durante la cual los espermatozoides son llevados al oviducto de manera sorpresiva desde los reservorios en el cérvix y la unión útero tubar. Estos sitios de colonización son los mismos lugares de depósito.

Sin embargo, investigaciones recientes
demuestran que los espermatozoides que llegan al
oviducto a los pocos minutos después de la cópula no
eran viables, por lo que su importancia puede solamente
representar un exacerbación de la actividad de transporte por la
contractilidad del tracto femenino junto con la cópula. Estudios recientes
demuestran que un alto porcentaje de los espermatozoides depositados en el
útero en la vaca se pierden por transporte retrógrado. En la IA el 60% se
pierden por este mecanismo dentro de las 12 horas siguientes a su depósito. Por
lo anterior se ha recomienda depositar el semen en lo más anterior posible del
cérvix, lo que implica una nueva instrucción del personal
responsable de la inseminación.
Luego del servicio en la vaca los espermatozoides deben sortear el sistema altamente circunvolucionado del cerviz por medio de su capacidad natatoria en el medio mucoso compuesto por sialomucinas y sulfomucinas. Las primeras muy viscosas desempeñan un mecanismo de lavado y las segundas más fluidas facilitan el movimiento y la natación del esperma.
Luego del servicio en la vaca los espermatozoides deben sortear el sistema altamente circunvolucionado del cerviz por medio de su capacidad natatoria en el medio mucoso compuesto por sialomucinas y sulfomucinas. Las primeras muy viscosas desempeñan un mecanismo de lavado y las segundas más fluidas facilitan el movimiento y la natación del esperma.
El tiempo requerido para que el espermatozoide
móvil entre y atraviese este medio especial influye significativamente en la
fase de transporte lento. (SENDER)
La producción diaria de espermatozoides y la
cantidad de espermatozoides fértiles está notablemente relacionada con el
tamaño testicular. Este se estima por la longitud y el grosor o bien por la
circunferencia escrotal. Sin embargo estos parámetros están influenciados por
la raza, el fenotipo y la edad, no pudiendose establecer estandares para todas
las razas y edades.
El bovino eyacula de 4 a 10 ml de semen con un
contenido espermático de 800 a 2.000 millones de espermatozoides por ml. Siendo
depositado en la parte anterior de la vagina. El total de espermatozoides por
eyaculado se incrementa con la edad, hasta los siete años y después decrece.
Existen grandes diferencias entre las
características de producción de espermatozoides entre toros, entre el primer y
segundo eyaculado y entre los intervalos de una recolección a otra.
La utilización prolongada de eyaculados
colectados a una frecuencia elevada, comenzando en la pubertad, no tiene efecto
negativo sobre el crecimiento o la eficiencia reproductiva de los toros. (HAFEZ)
MADURACION FINAL DEL OOCITO
La maduración del oocito
no se limita a la fase folicular, sino que
ocurre desde la fase de embriogénesis. Se
presenta en cuatro fases que son:
1.- División mitótica de las
CGP.
2.- Detención del desarrollo
nuclear en fase de dictioteno.
3.- Crecimiento citoplasmático.
4.- Reinicio de la meiosis.
La división mitótica se efectúa
antes del nacimiento y asegura que la
hembra nazca con una cantidad
suficiente de células germinales que formarán
el reservorio de los futuros folículos. La
última división mitótica de la oogonia se
constituye en un paso importante para que
el oocito primario entre en la primera división
de la profase meiótica. Durante este período
el núcleo del oocito primario permanece inactivo
hasta que es estimulado por las
gonadotropinas luego de la pubertad, hasta
la ovulación.
El
propósito de esta
suspensión es la de
inactivar el ADN
en el gameto
femenino de tal
manera que no
sea vulnerable a posibles
lesiones durante el
período de vida
de la hembra.
Las lesiones o daño del
gameto femenino puede comprometer la
reproducción al inducir la muerte
embrionaria después de la fecundación.
Una vez el folículo
entra en la fase de dominancia, el oocito
alcanza un tamaño mínimo crítico y adquiere la capacidad de
reiniciar la meiosis, cuando ocurre la descarga
preovulatoria de LH.
Poco después del pico de
LH, las uniones GAP entre las células de la
granulosa y el oocito se deterioran. Cuando las
proyecciones de las células de la granulosa
se disocian del citoplasma del oocito, el
AMPc junto con el inhibidor meiotico del oocito
dejan de suspender el desarrollo del oocito
y este queda libre para iniciar la
primera división meiótica.
En el folículo dominante, el
núcleo del oocito comienza a migrar hacia
la periferia y se adosa contra la membrana plasmática
del oocito, lo que se constituye en
el primer signo morfológico de la iniciación
de la maduración final del oocito.

La activación del ovocito,
expresada en el Reinicio
de la meiosis se puede poner en
evidencia por la eliminación del segundo polocito,
20 a 30 minutos después de la
fusión gamética. El set haploide de cromosomas
de ovocito forma el pronúcleo hembra
que junto con elpronúcleo macho restablece el
número diploide de cromosomas de la
especie para iniciar el desarrollo de
un nuevo individuo. (BARROS)

Después de la fecundación sucede la segunda división meiótica produciendo el segundo cuerpo polar. En algunos casos, el primer cuerpo polar se divide produciendo dos cuerpos polares hijos, por lo que se pueden ver tres cuerpos polares.
MADURACION DEL ESPERMATOZOIDE
El período de vida de los espermatocitos
secundarios es de unas pocas horas, dividiendose rápidamente por mitosis para
dar lugar a las espermátidas, también haploides para transformarse en
espermátidas maduras elongadas con cabeza y cola que posteriormente serán
liberadas en la luz del túbulo seminífero. Estos cambios tienen lugar en el
núcleo y el citoplasma. Estas células están unidas entre sí por
puentes citoplasmáticos y en comunicación con las células
nutricias o de Sertoli, las que inducen a partir de
células señalizadoras la maduración final de la
espermiogénesis.
La espermiogenesis consta de cuatro fases, las
cuales se clasifican mediante la reacción de PAS (Periodic Acid Shiff) al
teñir los componentes acrosómicos en desarrollo de un color rojo oscuro
El papel del espermatozoide
maduro es el de liberar el material genético a
un oocito maduro durante la fecundación. Para
que estas células sean capaces de fertilizar, las
espermátidas esféricas sufren una serie de cambios en los
cuales el núcleo se vuelve altamente condensado, se
forma el acrosoma y la célula se hace
móvil. La capacidad de nadar o motilidad requiere del
desarrollo de un flagelo y una fuente generadora
conocida como hélice mitocondrial.
Este proceso consta de
cuatro fases: Fase de Golgi – Fase de Capuchón – Fase de
Acrosomal – Fase de maduración

La Fase de Golgi es el primer paso para la formación del acrosoma. En esta fase la organela del mismo nombre se acerca al núcleo, desprende vesículas que se le sobreponen y poco a poco se unen para convertirse en la vesícula acrosomal que se localiza en la parte apical del núcleo. Los centríolos situados en forma de T, muy cercanos al aparato de Golgi, van migrando hacia lo que será la base del núcleo.
El centríolo proximal se sitúa en la parte basal
del núcleo y, a partir del centríolo distal crece el axonema conformado
por dos microtúbulos centrales y 9 pares de microtúbulos periféricos.

La
inactividad transcripcional del núcleo hace que el espermatozoide sea
dependiente de modificaciones postranscripcionales como la fosforilación de
proteínas necesarias para adaptar su función de acuerdo a las necesidades.

El acrosoma es una membrana lisosomal que contiene enzimas hidrolíticas como la acrosina, hialuronidasa, zona lisina, esterasas e hidrolasas ácidas, necesarias para la penetración de la zona pelúcida del oocito.

Finalmente se sucede la Fase de
Maduración, donde se observan
las características finales de los espermatozoides: forma de la cabeza
característica de cada especie (oval y plana), cubierta en sus dos terceras
partes por el acrosoma; y la cola compuesta por las piezas media, principal y
terminal; en la pieza media se encuentran las mitocondrias en forma de hélice.
En esta fase se elimina gran parte del citoplasma por desplazamiento del mismo
hacia la pieza terminal de la cola originando la llamada Gota Citoplasmática.
El proceso de maduración termina con
la Espermiación, o liberación de los
espermatozoides a la luz del túbulo seminífero. Mediante movimientos
peristálticos los espermas son transportados de la rete testis a los
ductos eferentes y de allí al epidídimo en cuya cola se almacenan.
En la pubertad y a lo largo de la vida adulta
del macho se producen continuamente en los túbulos seminíferos las células
germinales y la posterior liberación de espermatozoides en el proceso de
espermatogenesis, los cuales son reemplazados por espermatozoides de nuevas
generaciones. Cada nueva generación va empujando la anterior hacia el lumen del
túbulo seminífero, a tiempos e intervalos regulares lo que constituye el
llamado ciclo del epitelio seminífero, cuya duración de cada ciclo en el
toro es de 14 días y en el caballo 12 días.

Se requieren de 4 a 5
ciclos para que una espermatogonia tipo A del primer ciclo complete la
metamorfosis para un total de 60 días, duración de la espermatogénesis.
ESPERMATOZOIDE FERTIL
Para que el
espermatozoide sea capaz de fecundar el óvulo debe sufrir una serie de cambios
tanto en los testículos como en las vías genitales femeninas.
CAMBIOS
EN EL EPIDIDIMO
El espermatozoide una
vez ha sido expulsado del tubo
seminífero, no es apto para fecundar.
Comienza a adquirir ésta capacidad en
su paso por el epidídimo, en donde
sufre una serie de cambios morfológicos,
fisiológicos y bioquímicos cuyo conjunto
recibe el nombre de Maduración Epididimal
y tiene una duración de 10 a 15 días. Los
cambios en el epidídimo dependen de las secreciones del epidídimo y del tiempo
de transporte.
Estos cambios son:
- Adquisición de la motilidad (GADDUM)
- Modificaciones en la distribución y densidad de los grupoas aniónicos en su superficie (YANAGIMACHI)
- Formación de puentes de sulfuro -S-S- tanto en el núcleo como en estructuras de la cola del epidídimo (CHANG)
CAMBIOS EN LA VIA GENITAL FEMENINA
El conjunto de cambios señalados, es quizás lo que le permite al espermatozoide adquirir eventualmente, su capacidad fecundante.
El espermatozoide eyaculado, a
diferencia del testicular, es capaz de fecundar,
debido a los cambios que ocurren en
condiciones normales en la vía genital femenina.
Es necesaria la Capacitación
Espermática para la fecundación del oocito.

El espermatozoide adquiere la capacidad de mover el flagelo en su
tránsito por el epidídimo, pero el movimiento empieza después de la
eyaculación. Este proceso es conocido como la Activación del Esperma. (OLIVERA)
La movilidad del esperma se desencadena por cambios en el medio iónico
extracelular, por interacción con ligandos específicos y por glucosa, presentes
en el líquido seminal y en el tracto reproductivo femenino; estos cambios
inducen señales citosólicas flagelares, a través de la fosforilación de
proteínas, de canales de Ca++ y de vías dependientes de nucleótidos cíclicos
(GMPc y AMPc).
El espermatozoide, activado y capturado por las microvellosidades
del istmo del oviducto se capacita, y esto desencadena señales
intracelulares que inducen la hiperactivación. La activación y la
hiperactivación utilizan mecanismos moleculares similares para generar el
movimiento del flagelo cuyo eje funcional es el axonema y cuya proteína
motora principal es la dineína. El axonema, además, está compuesto por
microtúbulos, moléculas chaperonas, proteínas fijadoras de calcio y proteínas
quinasas/fosfatasas.

CAPACITACION ESPERMATICA
“La capacitación
espermática comprende todos
los cambios concurrentes
que permitan que
frente a niveles
fisiológicos de Ca ++
libre, se innduzca la “Reacción
Del Acrosoma” y que
el flagelo de
la cola sea “Hiperactivado” (BEDFORD)
Los espermatozoides son retenidos en las criptas oviductales y allí
pierden los factores decapacitantes como mucopolisacáridos y proteínas que
habían aportado las glándulas anexas; éste es el comienzo del proceso conocido
como Capacitación , nombre que indica el potencial que adquiere el
espermatozoide para hiperactivarse y para lograr la Reacción Acrosomal. Este
proceso se lleva a cabo en las criptas del istmo donde se adosan los espermas,
y termina con la liberación del mismo hacia el ámpula; aquí al encontrar el
oocito ocurre el reconocimiento y la adherencia para que el espermatozoide
empiece a atravesar la zona pelúcida.
La capacitación, se
caracteriza por la salida de colesterol de la membrana y el ingreso de Ca++ y
HCO3- al citosol (OLIVERA) lo cual tiende
a aumentar la fluidificación de la membrana,
niveles de AMPc y cambios en algunas enzimas
como la proteinKinasa (LANGLAIS)
Todos los espermatozoides no
son capacitados al mismo tiempo, sino que
lo hacen en un período relativamente largo
de varias horas dependiendo de su
localización en el tracto femenino
No hay duda que la membrana
plasmática del espermatozoide, particularmente la cabeza,
sufre una serie de cambios bioquímicos
durante la capacitación. Durante la mezcla del esperma
con el plasma seminal, este último recubre el esperma con
varias proteínas, las cuales son retiradas en
el tracto genital femenino.
Un concepto importante en relación
con la capacitación es el que este proceso puede
ser reversible si se reincorporan los espermatozoides capacitados
al plasma seminal. Por ejemplo, cuando se retiran espermatozoides
capacitados del tracto reproductivo femenino y se mezclan con
plasma seminal, estos se decapacitan nuevamente, necesitándose de un
nuevo período de capacitación en el tracto genital para que
puedan ser fértiles. (SENDER)
HIPERACTIVACION DE LA MOTILIDAD
La aparición de la Hiperactivación de la Motilidad
es necesaria para la penetración de
las cubiertas ovocitarias, particularmente la
Zona Pelúcida. (BEDFORD)
Cuando el espermatozoide
alcanza el istmo del oviducto inicia un movimiento asimétrico, amplio y
acelerado del flagelo (característico de la hiperactivación), lo que lo lleva a moverse en círculos y lo ayuda
a liberarse de las criptas oviductales para avanzar a través del lumen y
alcanzar el ámpula, atravesar el cúmulus ooforus (células de la granulosa que
rodean el oocito) y aponerse a la zona pelúcida donde es reconocido (reconocimiento
entre gametos)
La hiperactivación de
la motilidad sucede en la ámpula del
oviducto, varía de una motilidad progresiva en línea a
una motilidad acelerada en círculos y se cree es originada
por la presencia de algunas moléculas específicas
producidas en el epitelio de esta zona y se desencadena
por cambios en el medio iónico extracelular, por interacción con ligandos
específicos y por glucosa, presentes en el líquido seminal y en el tracto
reproductivo femenino; estos cambios inducen señales citosólicas flagelares, a
través de la fosforilación de proteínas, de canales de Ca++
y de vías dependientes de nucleótidos cíclicos
(GMPc y AMPc) (SENDER-
OLIVERA)
Los ligandos específicos más conocidos son: la progesterona y el
esteroide sulfatado SAAF (Sperm Activating and Attracting Factor) que inducen
la entrada de Ca++; el péptido activador de espermatozoide (PAS) y el péptido atrial
natriurético (PAN) que actúan, ya sea por medio de un receptor de membrana, o
por activación directa de la guanilil ciclasa ligada a membrana (GCm).
Otros ligandos específicos son los factores de tipo olfatorio y odorante
(hOR17-4) producidos por el oocito para inducir la quimiotaxis del
espermatozoide.
La hiperactivación depende
de la temperatura con un tiempo
mínimo de 2.5 Hs a 40° C
y de 12 Hs a 23° C. Normalmente
la hiperactivación tiende a ocurrir al
mismo tiempo que la reacción del
acrosoma. (MAHI)
La hiperactivación facilita el
contacto del espermatozoide con el oocito, mediante la interacción
de las proteínas de la superficie de la zona ZP
con las proteínas del acrosoma.
La ZP contiene tres
glicoproteínas denominadas Zona Proteinas 1-2-3 (ZP1-ZP2-ZP3). La ZP1 y ZP2
suministran las proteínas que conforman la estructura e integridad
de la ZP. La ZP3 actúa como un receptor hormonal y se une a
proteínas de la membrana del espermatozoide.
La membrana plasmática
del espermatozoide posee dos sitios de unión a la ZP. El primero
se denomina como Sitio de
Unión Primario de la ZP, responsable de la
adherencia del esperma a la ZP. El segundo se denomina Sitio de Unión Promoción de la Reacción del
Acrosoma. Cuando se produce el contacto de estos
sitios con la molécula ZP3 se produce una señal de transducción,
con lo cual se inicia la reacción del acrosoma
REACCION
DEL ACROSOMA.
La reacción del acrosoma comienza cuando la membrana plasmática del
espermatozoide se fusiona en múltiples sitios con la membrana
acrosomal externa, en donde se presenta un proceso de perforación o
fenestración y se forman pequeñas vesículas, proceso llamado vesiculación, que permite
la dispersión del contenido acrosomal y la
membrana nuclear del esperma queda rodeado por la membrana
acrosomal interna. La lesión de la membrana acrosomal y la
membrana plasmática es irreversible y se debe a los
cambios de presión osmótica, enfriamiento brusco, calentamiento
brusco o cambios marcados de pH. La liberación de las
enzimas acrosomales permite que el
espermatozoide digiera las membranas de la ZP y penetre
el oocito.

Austin y Bishop encontraron
espermatozoides cuyo acrosoma había reaccionado
mientras se movían entre las células del cúmulo ooforo. La
fenestración que se produce durante la reacción del acrosoma, permite la salida
gradual de hialuronidasa, cuya
localización intracrosómica ha sido comprobada, facilitando la
penetración del cúmulo por parte del espermio.
La reacción del acrosoma compromete
fundamentalmente la porción principal y más anterior del acrosoma, quedando la
porción posterior o segmento ecuatorial sin vesicularse.
(BARROS) La prematura reacción del acrosoma impide
la fertilidad del espermatozoide.
Experimentalmente se ha demostrado que la reacción del acrosoma es Ca ++
dependiente. Al incubar espermios en un medio sin
Ca ++ la reacción
no se presenta. Posteriormente al agregar Ca
++ éstos sufren la reacción del acrosoma y
son capaces de fusionarse con los ovocitos.
PENETRACION DEL ESPERMATOZOIDE
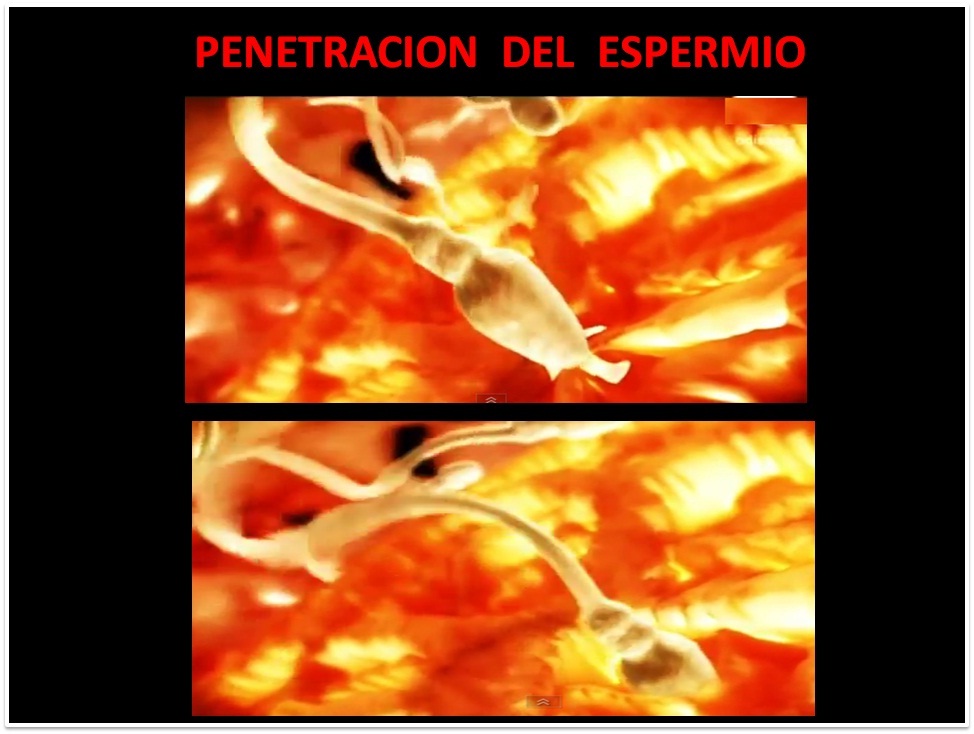
La fuerza
mecánica generada por
el movimiento flagelar
de la cola,
es suficiente para
empujar el espermio
a través de
un espacio oblicuo
de la ZP
denomiado Zona Lisina
FUSION DE LAS MEMBRANAS

.
La única región libre es
aquella que se encuentra por encima del sitio en donde se ubica la placa
de la segunda metafase. En ésta zona no se ha observado nunca fusión gamética.
Probablemente ésta diferenciación evita que el núcleo espermático sea eliminado
junto con el 2° polocito.

Durante
la primera y segunda división meiótica de la oogénesis, pequeñas
estructuras esféricas, densas denominadas Gránulos Corticales que
contienen mucopolisacáridos, proteasas, activadores de plasminógeno, fosfatasas
ácidas y peroxidasas, se mueven hacia la
periferia del citoplasma y entran en Exocitosis o
Decondensación lo cual permite la salida de
su contenido originando el Bloque o Reacción
de La ZP que impide la penetración de más
espermatozoides o Polispermia.
Después
de la penetración del espermatozoide tiene lugar la Decondensación
del Núcleo de tal manera que los cromosomas
del macho se aparean o fusionan con los
cromosomas del pronúcleo femenino, formándose por
lo tanto un solo núcleo con el ADN del macho y la
hembra, evento denominado Singamia que da lugar
a la formación del Zigoto como primer
paso para la embriogénesis.
REFERENCIAS
AUSTIN C.R. BISHOP M.W.H.
.- Role of the rodent acrosome and perforatorium infertilization. Proc.
Roy. Soc. Lond. Ser. B. 149: 241 1.958
.
BARROS C. .-Fecundación. Curso
Internacional relaciones Materno-Fetales y Nuevas Tecnologías
en Transferencia De Embriones. 169 – 202 1.984
BEDFORD JM. Significance of need
for sperm capacitation before fertilization
in Eutherian mammals. Biol. Reprod. 28: 108 –120
1.983
CHANG M.C..- Fertilizing
capacity of spermatozoa deposited into the
fallopian tubes. Nature – Lond – 1.951.
GADDUM P. .- Sperm
maturation in male reproductive tract:
Development of motility Anat. Rec. 161: 471 – 482 . 1.986
HAFEZ E.S.E. .-
Transporte y Supervivencia de los Gametos.
Reproducción e Inseminación Artificial en Animales. 182 – 204.
1.989
LANGLAIS J. KAN
FWK. GRANGER L. RAYMOND L BLEAU G. ROBERTS KD. Identification
of sterol receptors that stimulate choresterol effux from human
spermatozoa during in vitro capacitation. Gamet Res. 20: 185 – 201
1.998
MAHI C.A. y
YANAGIMACHI R. The efect of temperature, osmolarity and
hydrogen ion concentration on the activation and acrosome reaction
of golden hamster spermatozoa. J. Reprod. Fertil. 35: 55 – 66 1.973
MIL.- www.citrus.cc.ca.us/
RUIZ . – OLIVERA A. Ovarían follicular
dynamics in suckled o zebu cows (Bos Indicus) by real time
ultrasonography. Anim Reprod. Sci; 64:211-220.
SENGER P.L. PATH WAY TO PREGNANCY AND
PARTURITION Segunda Edición. Washington State Uniersity Research &
Technology Park 1610 NE Eastgate Blvd., Pullman, WA 99163-5607 Website:
YANAGIMACHI R.
.-Mechanisms of fertilization in mammals. En
: L. Mastroianni Jr, Bidggers
Eds.Fertilization and Embryonic Development in Vitro. Plenum Publishing
Co. pp 81 – 182

Comentarios
Publicar un comentario